Nombre científico:
Sardinops sagax (Jenyns, 1842) (Fig.1).
Otros nombres científicos utilizados:
Sardinops caeruleus (Girard, 1856), Sardinops sagax caeruleus, Sardinops caerulea.
Otros nombres comunes utilizados:
Sardina del Pacífico, sardina de California.
CARACTERÍSTICAS
Cuerpo algo cilíndrico, no muy comprimido. Color: dorso verde-azulado, flancos plateados, una serie de manchas oscuras a lo largo de la línea medio-lateral del cuerpo, y a veces, una segunda serie (o aún una tercera) más corta y más abajo (Whitehead y Rodríguez-Sánchez, 1995a).
Esta especie habita desde Alaska hasta el Golfo de California y en años fríos se distribuye hasta Mazatlán, Sinaloa (Murphy, 1966; Miller y Lea, 1972; Radovich, 1982; Whitehead, 1985); es una especie pelágica predominantemente costera, que forma cardúmenes, pero puede encontrarse hasta 160 km de la costa, al menos durante el periodo de desove, frente a California, E.E.U.U. (Whitehead y Rodríguez-Sánchez, 1995a).
Se ha propuesto que existen dos centros de distribución:
1) En el Golfo de California, alrededor de las grandes islas, y
2) Al oeste de la península de Baja California, frente a Punta Eugenia, por lo que a partir de esos centros de distribución la sardina monterrey se expande y se contrae obedeciendo señales ambientales (Lluch-Belda et al., 1995, 2003). Por otro lado, se reconoce la existencia de dos a tres stocks que responden de manera independiente (Félix-Uraga et al., 2005; Smith, 2005).
La sardina monterrey es omnívora facultativa, aunque muestra preferencia por el fitoplancton, ya que componen su dieta principalmente de diatomeas (Cadet y Berner, 1959; Kawasaki, 1983; Manrique y Molina, 2000). López-Martínez et al., (1999) reportan predominio de las diatomeas del género Navicula, Dinophysis y Planktoniella y copépodos, en particular Calanus sp.
En el área de Ensenada, la época reproductiva es en invierno-primavera, con el máximo entre enero y marzo, aunque se presentan pulsos importantes en el segundo semestre del año, en temperaturas superficiales del mar (TSM) entre 14.4 y 19.8°C (Cotero-Altamirano et al., 2011a) (Tabla 3). La longitud media de madurez (L50) está relacionada con la variabilidad ambiental y el comportamiento de la flota, por lo que las estimaciones anuales son muy variables, ya que va de 166 mm, en 2006, hasta 177 mm, en 2007 (Cotero – Altamirano y Valles-Ríos, 2007, 2008). La longitud media de madurez (L50) más reciente fue de 173 mm (Cotero-Altamirano et al., 2011b). Asimismo, la fecundidad promedio ha variado entre 13,285 y 20,367 ovocitos por desove (Tabla 3) (Cotero-Altamirano et al., 2009, 2011a).
En Bahía Magdalena, el desove se realiza principalmente en invierno, a temperaturas entre 19.8 y 21.0°C (Funes-Rodríguez et al., 2001); Melo-Barrera et al., (2010) reportan el máximo reproductivo entre diciembre y marzo, con un segundo pulso de menor intensidad en junio-julio (Tabla 3); Torres-Villegas et al., (2007) mencionan que este segundo máximo se presenta en ocasiones, por lo que la temporada de desove varía entre 4 y 8 meses. La longitud media de madurez (L50) fue de 135-165 mm en el periodo de 1982-1992 (Torres Villegas et al., 1995); Alvarez-Trasviña (2008) reporta para hembras 165.9 mm y para machos 146.4 mm.
Tabla 3.- Características biológicas de la sardina monterrey de Ensenada, Golfo de California y Bahía Magdalena
| Area de estudio | Longitud patrón promedio (mm) | Longevidad (años) | Fecundidad promedio (ovocitos/desove) | Reproducción (máximo) |
| Ensenada | 190 150 a 218 | 7 | 24,282 13,285 a 20,367 | ene-mar |
| Golfo de California | 167 | 7 | 19,913 | nov-feb |
| Bahía Magdalena | 161 | 7 | — | dic-mar |
En el Golfo de California, el periodo reproductivo se realiza a fines del otoño-invierno-primavera, con un máximo que se sitúa entre noviembre y febrero, en ocasiones con un breve desove en verano (Nevárez- Martínez, 1990; Cisneros-Mata et al., 1991, 1997; Martínez-Zavala et al., 2006) (Tabla 3); ya que la duración e intensidad de este proceso se relaciona con la temperatura del agua Nevárez-Martínez, 1990). El desove se realiza con mayor frecuencia en la costa de Sonora, aunque se pueden encontrar huevecillos en el centro del Golfo, la mayor proporción del desove ocurre a temperaturas de 19.9±1.9°C. (Hammann et al., 1998). La talla de primera reproducción de la sardina monterrey puede variar. Las tallas mínimas registradas en la década de los ochentas eran de 130-135 mm (Torres-Villegas et al., 1986; Cisneros-Mata, 1987). En contraste, en 1994, la talla mínima de reproducción fue de 120 mm (Cotero-Altamirano, CRIP Ensenada, com. pers), en 1996/1997 de 107 mm y en 1998/1999 de 130 mm. La longitud media de madurez (L50) en la década pasada fue de 146-150 mm (Torres-Villegas et al., 1986); mientras que en 1994 fue de 152 y 150 mm para hembras y machos respectivamente (Cotero-Altamirano, 1999), en 1998/1999 a 2002/2003 varió entre 147 a 166 mm (Martínez-Zavala et al., 2006), indicando una gran variabilidad en la longitud de reproducción. Por la maduración asincrónica de los ovocitos, esta sardina realiza desoves múltiples, por lo que en la época de reproducción puede desovar cada 15 días (Macewicz et al., 1996).
| Autor(es) | L∞ (mm) | K (años-1) | t0 (años) | Area de estudio |
| Holt (1960)* | 260.0 | 0.5 | – | California |
| Beverton (1963)* | 290.3 | 0.4-0.5 | – | California |
| García-Franco et al., (1995a)*** | 285.55 | 0.69 | -0.150 | Baja California |
| Félix-Uraga (1990)* | 170.0 a 183.0 | 0.82 a 1.73 | -0.0086 a -0.0179 | Bahía Magdalena |
| Molina y Pedrín (1976)** | 198.6 | 0.32 | -1.95 | Golfo de California |
| Méndez-Dasilveria (1987)* | 160.0 | 2.065 | -0.0082 | Golfo de California |
| Jiménez-Rodríguez (1991)* | 202.9 a 245.6 | 0.28 a 0.59 | -.021 a -1.66 | Golfo de California |
| Estrada et al., (1986)*** | 227.5 | 0.39 | -0.235 | Golfo de California |
| Cisneros-Mata et al., (1989)*** | 224.5 | 0.60 | -0.16 | Golfo de California |
| Cisneros-Mata et al., (1991)*** | 250.0 | 0.43 | -0.4014 | Golfo de California |
| Gallardo-Cabello et al., (1991)** | 197.95 | 0.3612 | -1.6324 | Sur del Golfo de California |
| Nevárez-Martínez et al., (1993)*** | 242.0 | 0.43 | -0.405 | Golfo de California |
| Cisneros-Mata et al., (1997a)*** | 246.0 a 247.0 | 0.54 | -0.30 | Golfo de California |
| De Anda-Montañez et al., (1999)***1 | 257 (LT) | 0.38 | -0.30 | Golfo de California |
| Martínez-Zavala et al., (2000)*** | 227.0 a 224.0 | 0.55 a 0.53 | -0.299 a -0.312 | Golfo de California |
| Nevárez-Martínez y Santos-Molina (2001) * | 198.7 | 0.463 | -1.74 | Golfo de California |
| Martínez-Zavala et al., (2006)*** | 216.5 a 222.5 | 0.55 a 0.53 | -0.306 a -0.316 | Golfo de California |
| Nevárez-Martínez y Santos-Molina (2008)* | 210.2 a 212.2 | 0.50 a 0.46 | -0.971 a -1.145 | Golfo de California |
*Estudios basados en la lectura de edad en otolitos
** Estudios basados en la lectura de edad en escamas 1 se utilizó la longitud total (LT)
*** Estudios basados en el análisis de frecuencia de tallas
| Autor(es) | L∞ (mm) | K (años-1) |
| Holt (1960)* | 260.0 | 0.5 |
| Beverton (1963)* | 290.3 | 0.4-0.5 |
| García-Franco et al., (1995a)*** | 285.55 | 0.69 |
| Félix-Uraga (1990)* | 170.0 a 183.0 | 0.82 a 1.73 |
| Molina y Pedrín (1976)** | 198.6 | 0.32 |
| Méndez-Dasilveria (1987)* | 160.0 | 2.065 |
| Jiménez-Rodríguez (1991)* | 202.9 a 245.6 | 0.28 a 0.59 |
| Estrada et al., (1986)*** | 227.5 | 0.39 |
| Cisneros-Mata et al., (1989)*** | 224.5 | 0.60 |
| Cisneros-Mata et al., (1991)*** | 250.0 | 0.43 |
| Gallardo-Cabello et al., (1991)** | 197.95 | 0.3612 |
| Nevárez-Martínez et al., (1993)*** | 242.0 | 0.43 |
| Cisneros-Mata et al., (1997a)*** | 246.0 a 247.0 | 0.54 |
| De Anda-Montañez et al., (1999)***1 | 257 (LT) | 0.38 |
| Martínez-Zavala et al., (2000)*** | 227.0 a 224.0 | 0.55 a 0.53 |
| Nevárez-Martínez y Santos-Molina (2001) * | 198.7 | 0.463 |
| Martínez-Zavala et al., (2006)*** | 216.5 a 222.5 | 0.55 a 0.53 |
| Nevárez-Martínez y Santos-Molina (2008)* | 210.2 a 212.2 | 0.50 a 0.46 |
| Autor(es) | t0 (años) | Area de estudio | ||
| Holt (1960)* | California | |||
| Beverton (1963)* | – | California | ||
| García-Franco et al., (1995a)*** | -0.150 | Baja California | ||
| Félix-Uraga (1990)* | -0.0086 a -0.0179 | Bahía Magdalena | ||
| Molina y Pedrín (1976)** | -1.95 | Golfo de California | ||
| Méndez-Dasilveria (1987)* | -0.0082 | Golfo de California | ||
| Jiménez-Rodríguez (1991)* | -.021 a -1.66 | Golfo de California | ||
| Estrada et al., (1986)*** | -0.235 | Golfo de California | ||
| Cisneros-Mata et al., (1989)*** | -0.16 | Golfo de California | ||
| Cisneros-Mata et al., (1991)*** | -0.4014 | Golfo de California | ||
| Gallardo-Cabello et al., (1991)** | -1.6324 | Sur del Golfo de California | ||
| Nevárez-Martínez et al., (1993)*** | -0.405 | Golfo de California | ||
| Cisneros-Mata et al., (1997a)*** | -0.30 | Golfo de California | ||
| De Anda-Montañez et al., (1999)***1 | -0.30 | Golfo de California | ||
| Martínez-Zavala et al., (2000)*** | -0.299 a -0.312 | Golfo de California | ||
| Nevárez-Martínez y Santos-Molina (2001) * | -1.74 | Golfo de California | ||
| Martínez-Zavala et al., (2006)*** | -0.306 a -0.316 | Golfo de California | ||
| Nevárez-Martínez y Santos-Molina (2008)* | -0.971 a -1.145 | Golfo de California |
*Estudios basados en la lectura de edad en otolitos
** Estudios basados en la lectura de edad en escamas 1 se utilizó la longitud total (LT)
*** Estudios basados en el análisis de frecuencia de tallas
Patrones de reclutamiento
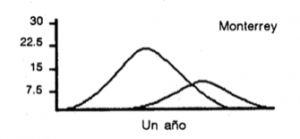
Se han analizado los patrones de reclutamiento pesquero de la sardina monterrey del Golfo de California (Cisneros-Mata et al., 1989, 1991), es decir, las épocas del año en que las especies se reclutan a la parte pescable, definida como la de menor vulnerabilidad, pero capturable (Sparre et al., 1989). En general, los resultados muestran que existen dos periodos de reclutamiento, siendo uno de mayor magnitud que el otro.
En la figura 2 se muestra de manera esquemática el patrón de reclutamiento de esta especie.